 |
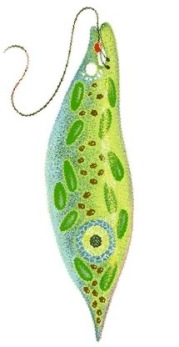
Das Augentierchen Euglena:ein einzelliger Geißelträger des Süßwassers
Domäne Eukaryota Supergruppe Excavata Stamm Euglenozoa oder Abteilung Euglenophyta Klasse Euglenoida oder Euglenophyceae Ordnung Euglenida | 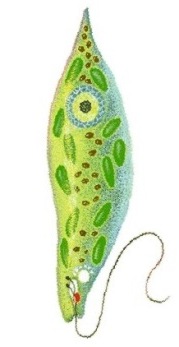 |
Wir stellen vor: Die Euglena-Verwandtschaft und ihre aktuelle systematische EinordnungDas Augentierchen Euglena und seine Verwandten, zu denen rund 40 Gattungen und 800 Arten zählen, sind einzellige Geißelträger (Flagellaten), die meist im Süßwasser in so großen Massen vorkommen, dass sie eine Wasserlache grün anzufärben vermögen. Die grüne Farbe deutet darauf hin, dass die meisten Arten den grünen Farbstoff Chlorophyll enthalten und mit seiner Hilfe Photosynthese betreiben, sich also in punkto autotropher Ernährung wie Pflanzen verhalten. Es gibt aber auch Formen, die wie die Tiere heterotroph von im Wasser gelösten Substanzen leben. Viele Arten pflegen sogar beide Ernährungsweisen gleichzeitig. (Hierzu weiter unten mehr.) Früher rechnete man daher die Eugleniden wahlweise zu den einzelligen Algen, genauer: zur Abteilung Euglenophyta, also zum Pflanzenreich, oder aber zur Klasse Flagellata (=Mastigophora) innerhalb der Protozoen, also zu den einzelligen Tieren. Danach gehörten sie eine Weile zum Stamm Sarcomastigophora, also zu den Einzellern mit Geißeln oder Pseudopodien, denn es hatte sich herausgestellt, dass manche Flagellaten so genannte Scheinfüßchen ausbilden und einige Amöben als Reaktion auf bestimmte Umwelteinflüsse Geißeln entstehen lassen können. Seit den achtziger Jahren des 20. Jahrhunderts stellte sie der Systematiker in ein eigenes Reich der Lebewesen, nämlich ins Reich der Protoctista, vormals Protista (der Einzeller mit echtem Zellkern). Für moderne Systematiker zählen allerdings die vielen Unterschiede der Eugleniden zu allen anderen Protoctisten so stark, dass sie geneigt sind, innerhalb der neu gegründeten "Supergruppe" Excavata ein Taxon Euglenozoa oder Euglenophyta für sie einzuführen, zu dem nicht nur die Eugleniden, sondern auch die mit ihnen verwandten, ebenfalls einzelligen, aber parasitisch lebenden Trypanosomen gerechnet werden. [Worüber sich Fachleute so alles streiten können ...]
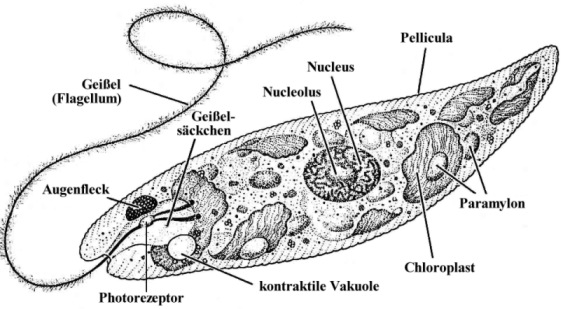
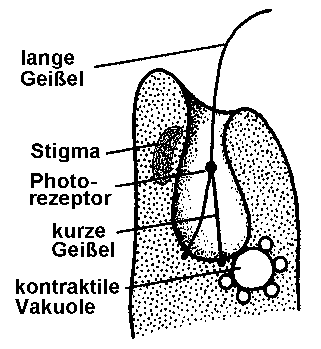
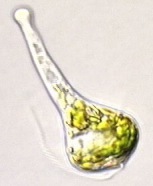
Besonderheiten der Zellmembran und der Begeißelung von EuglenaEuglena besitzt wie die anderen Vertreter der Eugleniden einen schraubenförmig verdrehten Zellkörper. Die Zellmembran liegt in Form einer festen, aber biegsamen, aus Proteinen aufgebauten Pellicula vor, die an der Oberfläche enge, feine, meist spiralig umlaufende Streifen ausbildet. Genauere Untersuchungen haben gezeigt, dass dicht unterhalb der Zellmembran liegende, an Membranproteinen hängende "epiplasmatische Proteine" ein Membranskelett formen, das mit den Zisternen des Endoplasmatischen Reticulums und mit Mikrotubuli in Verbindung steht. Im Unterschied zu anderen einzelligen, ebenfalls grün gefärbten Algen ist Euglena also von keiner Zellwand aus Cellulose umgeben. Am Vorderende entspringen 1 bis 2 Geißeln (Flagellen; von lat. flagellum = Peitsche) einer säckchenartigen Vertiefung, die man als Geißelsäckchen, Ampulle oder - vor allem im anglo-amerikanischen Schrifttum - als Reservoir (oder "flagellar pocket") bezeichnet. [Erstaunlich, was für merkwürdige Säckchen es gibt.] Es können bei den Eugleniden zwei lange, "behaarte" (d.h. mit vielen kleinen abstehenden Mastigonemen besetzte) Geißeln vorhanden sein oder - wie bei den meisten Arten der Gattung Euglena - nur ein langes, nach vorne gerichtetes und der Fortbewegung dienendes Flagellum, das man auch Undulipodium (von lat. undulare = sich schlängeln; podium = fußartiger Fortsatz) nennt, wobei dann aber noch eine kurze Geißel im Geißelsäckchen liegt, die man als stark reduzierte zweite Geißel ansehen kann. Je ein Paraxonemalstab flankiert - als typisches Sondermerkmal der Euglenozoa - das Axonem sowohl der langen als auch der kurzen Geißel.Die kontraktile Vakuole pumpt überschüssiges Wasser aus der ZelleIn das Geißelsäckchen wird auch die kontraktile Vakuole entleert, die das in die Zelle eingedrungene Wasser entfernt. Es handelt sich dabei um ein pulsierendes Bläschen, das sich rhythmisch mit Wasser füllt, wobei sich das im Zellleib angereicherte überschüssige Wasser zunächst in umliegenden kleineren Bläschen sammelt, um von dort aus in die kontraktile Vakuole geleitet zu werden, von wo es periodisch nach außen abgegeben wird. [Wenn Euglena das nicht laufend täte, würde die arme Zelle ganz schön schnell anschwellen und platzen. *Plop!* Und das wär's dann gewesen ...] Die wenigen im Meer lebenden Eugleniden benötigen eine solche Einrichtung nicht, weil der viel höhere Salzgehalt des Meerwassers nach den osmotischen Gesetzen dafür sorgt, dass kein Wasser in den Zellkörper eindringt.
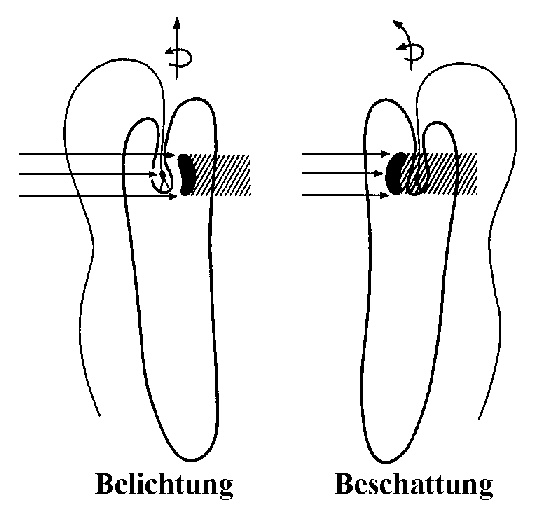
Unglaublich: Euglena kann sehen! - Die phototaktische Ausrichtung zum LichtNoch innerhalb des Geißelsäckchens ist am Anfangsabschnitt des Undulipodiums eine Schwellung zu erkennen, der so genannte Paraflagellarkörper. Dieser dient als Photorezeptor der Aufnahme von Lichtreizen und ermöglicht es Euglena, phototaktisch zu reagieren. Für die Lichtabsorption scheinen dabei Pterine und Flavine zuständig zu sein, wobei die Pterine als Antennenpigmente fungieren, die die Lichtenergie zu den Flavinen, den eigentlichen Rezeptorpigmenten, weiterleiten. Zusätzlich liegt von der Plasmaseite aus ein roter Augenfleck, das so genannte Stigma, der Membran des Geißelsäckchens seitlich an. Es handelt sich dabei um eine Ansammlung von Lipidtröpfchen, die - jeweils von einer Biomembran umhüllt - massenhaft Pigmente, vor allem das rötliche Carotinoid Astaxanthin, enthalten. Der Augenfleck nimmt aber selbst keine Lichtreize auf, hat also gar keine Augenfunktion, wie man früher annahm, sondern wirkt wie eine Art Sonnenschirm, der den Paraflagellarkörper der Geißel beschattet, so dass diesen kein seitlich einfallendes Licht treffen kann. [Wer hätte das gedacht: ein Sonnenschirm innerhalb einer winzigen Zelle!] Da sich Euglena bei der Fortbewegung um die Längsachse dreht, wird bei seitlicher Einfallsrichtung der Sonnenstrahlen der lichtempfindliche Teil der Geißel abwechselnd beschattet und belichtet, denn dann gerät der das Licht auffangende "Schirm" immer wieder zwischen die Lichtquelle und den Photorezeptor.
Es können also nur direkt von vorn einfallende Lichtstrahlen den Photorezeptor so erregen, dass die Euglena-Zelle einen dauerhaften, ununterbrochenen Lichtreiz empfängt. Sie wird ihren Zellkörper also durch Änderung des Geißelschlags nach und nach so ausrichten, dass es zu keiner rhythmischen Beschattung des Paraflagellarkörpers mehr kommt. Nach dieser Kurskorrektur reagiert Euglena positiv phototaktisch und schwimmt bevorzugt in Richtung des Lichteinfalls. Dies jedoch nur bei gemäßigter Einstrahlung. Bei sehr hoher Lichtintensität weicht Euglena rückwärts schwimmend vor der starken Lichtquelle zurück, verhält sich also negativ phototaktisch. Dies ist kein Wunder, würde doch eine allzu intensive Belichtung die lichtempfindlichen Moleküle im Paraflagellarkörper zerstören. Am empfindlichsten reagiert der Photorezeptor, wenn blaugrünes Licht einer Wellenlänge von 495 Nanometern auf ihn trifft. Dies dürfte eine Anpassung an das Wasserleben sein, denn mit steigender Wassertiefe verengt sich das Lichtsprektrum immer mehr zum Blaugrünbereich hin. Im Zusammenspiel mit dem Augenfleck ermöglicht der Paraflagellarkörper also ein einfaches Richtungssehen. Fehlt der Photorezeptor, weil z.B. die Flavine zerstört wurden, reagiert Euglena nicht mehr auf Lichtreize. [Dann ist es endgültig aus mit dem Traum vom einzelligen Sehen!]
Gravitaxis: Euglena nutzt auch die Erdschwerkraft als OrientierungshilfeNeben der phototaktischen Ausrichtung nach dem Lichteinfall ist bei Euglena auch eine negative Gravitaxis beschrieben worden, die dafür sorgt, dass die Zellen sich entgegen der Erdschwerkraft in Richtung Wasseroberfläche orientieren, also nicht zu tief in dunkle Wasserbereiche absinken, in denen kaum noch Photosynthese betrieben werden kann. Interessant ist dabei, wie Euglena die Richtung der Schwerkraft ermittelt: Der Zellleib erweist sich als schwerer als das umgebende Medium und wirkt wie ein riesiger Statolith, der auf die unten liegende Membran drückt. [Ein ziemlich gewichtiger Brocken, so ein Zellleib, wenn Euglena damit seine Schwere feststellen kann, und das gänzlich ohne Waage!] Geschieht dies, werden winzige Kanäle in der Membran geöffnet, welche Calciumionen in die Zelle einströmen lassen und dadurch das Membranpotential ändern. Diese Änderung des elektrischen Membranpotentials signalisiert dann der Zelle, dass sie ihre Bewegungsrichtung zu ändern hat. [Erstaunlich, wobei elektrische Signale so alles eine Rolle spielen.] Zusammen mit der Phototaxis sorgt die Gravitaxis für eine hinsichtlich der Lichtausnutzung optimale Position der Euglenen im Wasser. Bei zu intensiver Bestrahlung mit ultraviolettem oder blauem Licht verliert Euglena allerdings dieses negativ gravitaktische Verhalten und reagiert positiv gravitaktisch, schwimmt also in Richtung der Erdschwerkraft von der Wasseroberfläche weg. Diese Verhaltensänderung stellt wohl eine Art Notsicherung für den üblen Fall dar, dass die Pigmente des Photorezeptors bei zu starker Sonneneinstrahlung zerstört wurden, denn in diesem Fall sollten die Euglenen nicht weiter auf die zerstörerische Lichtquelle zuschwimmen, sondern sich in die dunkleren Gefilde des tieferen Wassers zurückziehen, um nicht noch mehr vom grellen Licht geschädigt zu werden. [Dann heißt es: nichts wie weg von der Strahlenquelle!]
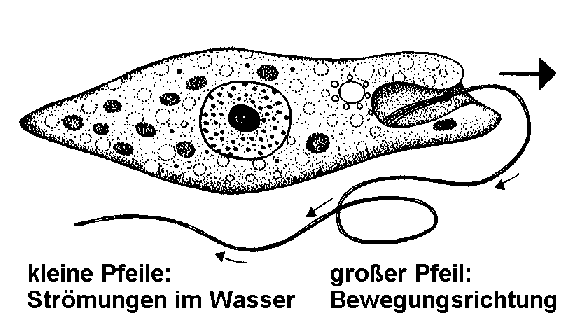
Die Fortbewegung mit Hilfe der SchwimmgeißelEs ist nicht ganz leicht, die Bewegungsweise von Euglena zu studieren, weil man zum einen die Schwimmgeißel, das Undulipodium, bei normaler Beleuchtung im Lichtmikroskop nur bei stärkster Vergrößerung und dann oft auch nur mehr schlecht als recht sieht, zum andern die Bewegungen meist sehr schnell erfolgen. Einige erfahrene Mikroskopiker raten daher, nicht nur spezielle Beleuchtungstechniken, etwa die Dunkelfeldbeobachtung, zu verwenden, sondern auch das Wasser, in dem Euglena schwimmt, mit etwas Agar, Gelatine oder Methylcellulose-Kleister zu versetzen, um die Bewegungsgeschwindigkeit der kleinen Zellen zu verringern. Auch die Abkühlung des Objektträgers mit Hilfe von Eiswürfeln soll bereits gute Dienste bei der Verminderung der Geschwindigkeit geleistet haben. [Klar: je kälter es ist, desto langsamer die Euglenen, denn nach der RGT-Regel verlangsamen sich in saukalter Umgebung alle Stoffwechselreaktionen. Und Muskelzittern bei eisiger Kälte kennen die Einzeller noch nicht!] Spezialisten setzen auch ein Stroboskop oder die Hochfrequenz-Blitzlicht-Kinematographie mit anschließender Einzelbildanalyse sowie in jüngerer Zeit die ausgefeiltesten digitalen Tricks ein, um die Bewegungsabläufe genauer zu studieren. An günstigen Objekten kann man dann beobachten, wie Euglena mit ihrer zunächst nach vorn in Schwimmrichtung aus dem Geißelsäckchen herausragenden, dann aber nach hinten umbiegenden Zuggeißel durch ein wellenförmiges und quasi nach Schiffsschraubenart schraubig-rotierendes Schlagen im dreidimensionalen Raum sich selbst nach vorne zieht, wobei sich gleichzeitig der Zellleib um seine Längsachse dreht. Teilweise erfolgt die Geißelbewegung aber auch nach dem Ruderprinzip schlagend-peitschend. Es gibt da viele Übergänge und unterschiedliche, schwer zu erfassende Bewegungsmuster. So kann Euglena mit einem "Spezialschlagprogramm" die Geißel vorübergehend auch nach der Seite schlagen lassen. Dieses Programm wird auf bestimmte Außenreize hin aktiviert. Die frühere Annahme, dass die Geißel in relativ gestrecktem Zustand eine kegelförmige Rotationsbewegung vollführt und dabei einen Sog erzeugt, der die Zelle vorwärts treibt, scheint sich nicht bestätigt zu haben. Bei Euglena viridis wurden mindestens zehn Geißelschläge pro Sekunde gemessen. Damit werden Geschwindigkeiten von ca. 0,15 mm pro Sekunde erzielt, was immerhin einem Vorwärtskommen um drei Körperlängen entspricht. [Eine Wahnsinnsgeschwindigkeit also.]
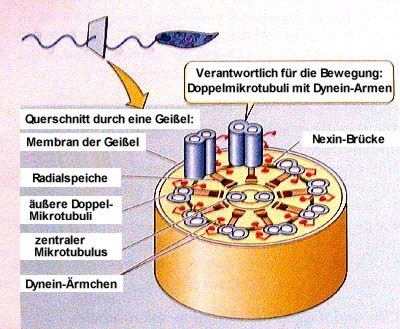
Der molekulare Mechanismus der Geißelbewegung ist dabei hochkompliziert. Nur soviel sei hier angedeutet: Elektronenmikroskopische Querschnittsbilder offenbaren, dass das 0,2 Mikrometer breite "Innenleben" der Geißel, das Axonem, ein typisches 9+2-Muster so genannter Mikrotubuli aufweist. Das heißt nichts anderes, als dass ein Undulipodium (das eigentlich nur eine sehr schmale, überlange, innen besonders strukturierte Ausstülpung der Zellmembran ist) der Länge nach von 20 winzigen Eiweißröhren, nämlich von 9 kreisförmig angeordneten Doppel-Mikrotubuli und zwei weiteren, zentral angeordneten, einzelnen Mikrotubuli durchzogen wird. Radialspeichen verbinden dann noch die äußeren Doppelröhrchen mit den beiden zentralen Mikrotubuli. Die Wandungen all dieser winzigen Röhren wiederum bestehen aus α- und β-Tubulin - kleinen, kugelförmigen Proteinen. Doch das ist noch längst nicht alles: Die Doppel-Mikrotubuli tragen an der einen Seite ihrer Längsachse in Abständen von ca. 17 Nanometern eine Vielzahl von paarigen Protein-Ärmchen aus Dynein, die dazu dienen, sich unter ATP-Verbrauch an den benachbarten Doppel-Mikrotubuli in Tausendfüßler-Manier quasi entlangzuhangeln. Wenn dieses Entlanghangeln mit Hilfe der Dynein-Ärmchen nur bei den Doppel-Mikrotubuli auf der einen, nicht aber bei denen auf der anderen Seite des Undulipodiums erfolgt, führt dies zur Krümmung der Geißel, zumal die Doppel-Miktrotubuli über die Radialspeichen mit der Geißelachse in stabiler Verbindung stehen und sämtliche Mikrotubuli an ihrem Ursprungsort, dem Basalkörper der Geißel, fest verankert sind. [Kompliziert, kompliziert! Wenden wir uns daher lieber anderen Themen zu.]
Lappenförmige Chloroplasten zur PhotosyntheseDie Zellen der verschiedenen Euglena-Arten enthalten ein bis zwölf Chloroplasten, die scheiben- bis lappenförmig aussehen können und nicht wie sonst üblich von zwei, sondern von drei Hüllmembranen umgeben sind. Das hat zu etlichen Spekulationen Anlass gegeben, etwa der, dass eine chloroplastenlose Ur-Euglena-Zelle endosymbiontisch kleine, einzellige, eukaryotische Grünalgen aufgenommen haben könnte [*mampf* *mampf* wie bei der Phagocytose], diese aber nicht wie die anderen üblichen Nahrungsteilchen vollständig verdaut, sondern nur teilweise abgebaut hat, so dass heute noch die Grünalgen-Chloroplasten - in einer Art Nahrungsvakuole (nämlich der dritten, äußeren Euglena-Chloroplastenmembran) eingeschlossen - als Reste der importierten Algenzellen weiterexistieren. Man nennt dieses Phänomen auch sekundäre Endocytobiose. [Die armen Chloroplasten der damals verschluckten Grünalgenzellen! Unverdaut liegen sie heute noch in den Euglenen herum und müssen bis in unsere Tage hinein für sie photosynthetische Frondienste leisten!] In den Euglena-Chloroplasten sind zahlreiche Pigmente enthalten, vor allem Chlorophyll a und b, aber neben β-Carotin auch die Carotinoide Diadinoxanthin, Neoxanthin und einige andere Farbstoffe, die man sonst nirgendwo in den grünen Pflanzen findet, was schon allein auf die Sonderstellung der Eugleniden im Reich der Protisten hindeutet. Mit Hilfe der Chlorophylle kann Euglena Photosynthese betreiben. Im Gegensatz zu den Grünalgen und höheren Pflanzen enthalten die Eugleniden aber vergleichsweise weniger Chlorophyll b. Und als Photosyntheseprodukt wird nicht Stärke, sondern Paramylon gebildet, das seiner chemischen Struktur nach β-1,3-Glucan ist, also ein Polysaccharid aus linearen Glucoseketten, die β-1,3-glykosidisch verknüpft sind. Dieses Reservekohlenhydrat lässt sich übrigens nicht wie Stärke mit Jodkalium-Jodid nachweisen. Seine Bausteine entstehen in einer bestimmten, für die Kohlendioxid-Fixierung und die Glucoseproduktion genutzten Chloroplastenregion, dem Pyrenoid. Das eigentliche Polysaccharid wird dann im Zellinneren in Form von Paramylon-Körnern abgelagert, die alle in der Mitte ein Loch aufweisen und von einer Membran umgeben im Cytoplasma liegen. In die Membran integriert ist ein Enzymkomplex, der für die Synthese und Anreicherung der Paramylon-Makromoleküle sorgt. |
 |
 |
 |
 |
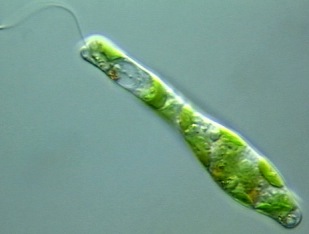
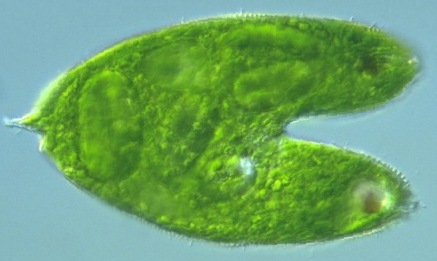
Im Mikroskop gut zu sehen: Bauchige Formveränderungen, die den Zellleib entlangwandernDer Zellkörper von Euglena ist biegsam und ermöglicht verschiedene Formveränderungen. Oft lässt sich im Mikroskop verfolgen, wie eine bauchige Erweiterung den ganzen Zellleib von vorne nach hinten durchläuft. Der Fachmann spricht hier von Metabolie oder einer metabolischen Bewegung. [Man stelle sich nur mal vor, der Mensch könnte seinen Bierbauch ebenfalls von oben nach unten am Rumpf entlangwandern lassen!] Verantwortlich für diese peristaltischen Verformungen entlang der Körperlängsachse sollen die unter der Pellicula liegenden Mikrotubuli sowie Kontraktionen der Actin- und Myosin-Filamente im Cytoplasma sein. Zwischendurch kann Euglena sogar eine fast kugelförmige Gestalt annehmen. [Wie manche pfundige Mitmenschen auch...]
Auch die heterotrophe Ernährungsweise kommt vorViele Eugleniden sind autotroph, d.h., sie können mit Hilfe ihrer Chloroplasten das Sonnenlicht zur Energiegewinnung und zum Aufbau organischer Substanz nutzen. Es gibt aber auch Formen, die gar keine Chloroplasten besitzen und sich rein heterotroph ernähren, indem sie sich andere kleine Einzeller einverleiben. Zur Nahrungsaufnahme dient ihnen ihr Zellmund, der in der Nähe des Geißelsäckchens liegt und dessen Mundapparat stabartige Strukturen aus Mikrotubuli besitzt. Besonders deutlich tritt dieses "Rod-organ" (wie es die anglo-amerikanische Forschung nennt) bei der Gattung Peranema in Erscheinung. Neben diesen "phagotrophen" Eugleniden gibt es auch noch "osmotrophe" Gattungen wie Distigma, die im Wasser gelöste Nahrungspartikel einfach über die gesamte Zelloberfläche aufnehmen.Wenn Euglena-Kulturen mit Zellen, die normalerweise Chloroplasten enthalten, lange im Dunkeln gehalten werden, stellt man fest, dass sie zu vollständig heterotropher Ernährung übergehen und die Chloroplasten zu degenerieren beginnen. Dann nehmen sie mit Hilfe der Endocytose gelöste organische Substanzen aus dem Umgebungsmedium auf, um aus diesen Energie zu gewinnen oder ihre Körpersubstanz aufzubauen. Dabei hat sich herausgestellt, dass zahlreiche Eugleniden in der Lage sind, energiereiche ATP-Moleküle nicht nur über die normale Zellatmung, d.h. unter Sauerstoffverbrauch, zu erzeugen, sondern unter ungünstigen Sauerstoffverhältnissen, ja sogar unter gänzlich anaeroben Bedingungen wochenlang eine Wachsesterfermentation zur Energiegewinnung durchzuführen. Für Spezialisten seien hier die wesentlichen Schritte dieses Prozesses kurz dargestellt ["Normalverbraucher" bitte jetzt weghören!]: Zunächst werden die Paramylonkörner zu Brenztraubensäure abgebaut und diese mittels eines speziellen, sauerstoffhassenden Enzyms, der Pyruvat:NADP+-Oxidoreduktase, zu Acetyl-CoenzymA decarboxyliert, welches dann in den Euglena-Mitochondrien unter anaeroben Bedingungen zur Fettsäuresynthese genutzt wird. Die entstehenden Fettsäuren nehmen dabei jene Elektronen und Protonen (in diesem Fall vom NADPH) auf, die bei der normalen Zellatmung im Rahmen der "biologischen Knallgasreaktion" letztlich der Sauerstoff empfangen würde. [Aber den gibt's ja unter anaeroben Bedingungen nicht.] Die Fettsäuren wiederum werden in Wachsester umgewandelt, die Euglena solange im parakristallinen Zustand als "Müll" im Cytoplasma ablagert, bis wieder normale Sauerstoffverhältnisse herrschen, unter denen die Wachsester oxidativ abgebaut und gewinnbringend entsorgt werden. [Wieder mal höchst kompliziert, das Ganze!]Wie dem auch sei: Im Inneren der Euglena-Zelle bleiben meist farblose Proplastiden zurück, die sich wieder zu normalen grünen Chloroplasten ausdifferenzieren können, wenn man die Euglena-Kulturen erneut und dauerhaft dem Licht aussetzt. [So schnell kann man aus farblosen Euglenen wieder farbige Zeitgenossen machen! Nicht auszudenken, wenn das bei langweiligen Menschen auch so schnell ginge! Immerhin deuten einige Experimente darauf hin, dass man mit intensivem Licht menschliche Depressionen vertreiben kann. Also schon mal ein Lichtblick! Und findige Geschäftsleute stellen bereits die dazu nötigen Leuchten her - für Leute, die das Tageslicht scheuen. Zum Glück kennen unsere Augentierchen noch keine Depressionen!] Lediglich dann, wenn sich unsere Euglenen etwas zu flott teilen und es dadurch mit der gerechten Verteilung der Zellorganellen hapert, können Zellen entstehen, die gar keine Chloroplasten mehr enthalten. Und die sind natürlich nicht mehr in der Lage, Photosynthese zu betreiben.
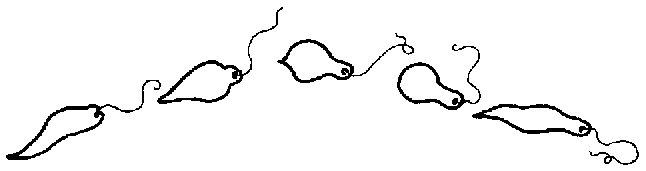
Euglena lebt asexuell! - Die ungeschlechtliche Vermehrung durch LängsteilungDie Vermehrung von Euglena erfolgt ungeschlechtlich und ausschließlich durch Längsteilung der Zelle in zwei Tochterzellen. Der gesamte Vorgang dauert etwa 2 bis 4 Stunden. Der Durchschnürung des Zellleibes geht wie üblich eine Kernteilung voraus. Interessant ist aber, dass bei den Eugleniden während der Mitose die Kernhülle erhalten bleibt. Spindelfasern und deutlich voneinander differenzierte Chromosomen sind in ihr kaum erkennbar. Und auch die sonst typische Äquatorialplatte, von der aus alle Chromosomen etwa gleichzeitig nach zwei Seiten auseinandergezogen werden, fehlt Euglena. Stattdessen sieht man Chromatingranula einzeln zu den Kernpolen wandern. Neben dem Kern verdoppeln sich auch andere wichtige Strukturen wie die Basalkörper der Geißeln oder das Stigma. Die Chloroplasten und Paramylon-Körner werden zu etwa gleichen Anteilen auf die Tochterzellen verteilt. Bei der eigentlichen Zellteilung beginnt sich der Zellleib am Geißelpol einzuschnüren, und im weiteren Teilungsverlauf vertieft sich der Spalt zwischen den beiden Zellhälften entlang der Längsachse der Zelle immer mehr, bis sie sich vollständig durchgeschnürt hat.
Die Teilung kann im beweglichen (s. Abb. oben) oder im unbeweglichen Zustand erfolgen. Letzteres geschieht, nachdem sich der Zellkörper der Mutterzelle vergrößert, die Geißeln abgeworfen und sich abgekugelt hat. Meist umgibt sich die Zelle dann mit einer Gallerthülle, wobei Schleimbläschen, die dicht unter den Streifen der Pellicula liegen, das gallertige Material nach außen abscheiden. [Das ist ein ähnlich glibberiges Material wie bei der Gelatine oder beim Wackelpudding. Und da drin liegen die Euglenen bei der Vermehrung? Igittigitt!] Unter günstigen Bedingungen kann es zu wiederholten Teilungen kommen, so dass zahlreiche Tochterzellen in der Gallerthülle liegen (so genanntes Palmella-Stadium). Nach Beendigung der Teilungsphase entstehen wieder neue Geißeln, und die Zellen schwärmen in die Umgebung aus.
Einige bekannte Euglena-ArtenEuglena viridis ist eine häufig in kleinen Tümpeln mit verunreinigtem Wasser anzutreffende Art. Sie wurde erstmals 1838 von Christian Gottfried Ehrenberg beschrieben, wird 40 bis 60 Mikrometer groß, besitzt eine spindelförmige Gestalt und verändert oft ihre Form. Die bandförmigen Chloroplasten in ihrem Inneren scheinen von einem Mittelpunkt zu entspringen. Euglena gracilis, eine weitere bekannte Augentierchen-Art, die mit 143.170 Basenpaaren das kleinste Genom eines funktionstüchtigen Chloroplasten enthält, wurde 2010 von der Deutschen Gesellschaft für Protozoologie sogar zum Einzeller des Jahres gewählt! Weil diese Art bereits erfolgreich als Sauerstofflieferant in bioregenerativen Lebenserhaltungssystemen eingesetzt wurde, ist sie auch für die Weltraumforschung interessant. Andere Arten sind z.B. Euglena spirogyra (neuerdings auch unter dem Namen Lepocinclis spirogyroides geführt), die durch ihre dicke, gelbliche Pellicula auffällt, an deren Außenseite zusätzlich spiralige Reihen von Warzen zu sehen sind, Euglena oxyuris (oder Lepocinclis oxyuris), die eine Länge von bis zu 530 Mikrometern erreicht, sowie Euglena mutabilis, die in Mooren und Tümpeln mit saurem Wasser vorkommt, nur eine sehr kurze Geißel besitzt und sich kriechend fortbewegt. Es gibt aber noch eine Vielzahl weiterer Euglena-Arten, von den zahlreichen anderen Gattungen der Eugleniden ganz zu schweigen.Weitere Infos:Andere Webseiten über Euglena:
Webseiten zu anderen Themen: |

| Autor: BKry | →Fragen? →Anregungen? →Kommentare?
|
Letzte Änderung: 15. Februar 2023 |