Die Fettsäuren
Einleitung
Fette sind sehr energiereich; sie liefern pro Gewichtseinheit mehr als doppelt soviel Energie wie Kohlenhydrate oder Proteine. Fette sind deshalb wichtiger Energieträger der Nahrung.Über den Normalbedarf aufgenommene Nahrungsbestandteile werden in Fette umgewandelt und in speziellen Geweben abgelagert, um bei Nahrungsmangel abgebaut zu werden. Fette stellen für den Menschen sowie viele Tiere und Pflanzen das wichtigste Speichermedium für Stoffwechselenergie dar. Bedeutsamer ist jedoch ihre Funktion als Reservestoff. Dabei führen Fette wegen ihrer Wasserunlöslichkeit nicht zu osmotischer Belastung. Für Tiere ist das Fettgewebe der Unterhaut zugleich ein thermischer Schutz. Die Rückgewinnung der gespeicherten Energie bei Nahrungsmangel erfolgt zumeist über die sogenannte β-Oxidation in den Mitochondrien (Karlson 1984).
In allen Geweben sind Fettsäuren außerdem wichtiger Bestandteil der Membranen. Deren physikalischen und physiologischen Eigenschaften werden zum großen Teil durch die Struktur der enthaltenen Fettsäuren bestimmt. Die Rate des oxidativen Abbaus von Fettsäuren entscheidet über deren Gehalt und (mittels selektivem Abbau) über das Fettsäureprofil eines Gewebes.
Die hier dargestellten Erkenntnisse stammen aus neueren Veröffentlichungen (1996); sie sind zumeist durch Versuche mit Nagetieren gewonnen worden.
Struktur und Nomenklatur
Die natürlich vorkommenden Fettsäuren enthalten fast alle eine gerade Anzahl von Kohlenstoffatomen. Am häufigsten sind die Säuren mit 16 bzw. 18 Kohlenstoffatomen; Palmitinsäure bzw. Stearinsäure. Die Struktur der Palmitinsäure ist exemplarisch in Abbildung 1 dargestellt. Viele Fettsäuren sind besser unter ihrem Trivialnamen bekannt als unter der Bezeichnung nach der systematischen Nomenklatur. Diese wird von dem Kohlenwasserstoff abgeleitet, der die gleiche Anzahl von Kohlenstoffatomen hat. Die korrekte Bezeichnung für Palmitinsäure lautet demnach n-Hexadekansäure.
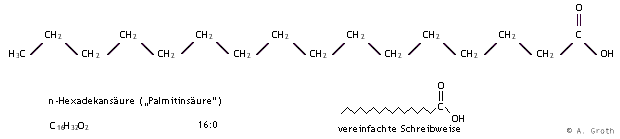
Abb. 1: Schreibweisen von Fettsäuren am Beispiel der Palmitinsäure (n-Hexadekansäure).
Fettsäuremoleküle können eine oder mehrere Doppelbindungen enthalten; man spricht dann von ungesättigten Fettsäuren. Die Doppelbindungen liegen fast ausnahmslos in der cis-Konfiguration vor. Mehrere Doppelbindungen sind meist durch eine CH2-Gruppe getrennt. Bei der Bezeichnung ungesättigter Fettsäuren werden die Positionen der Doppelbindungen von dem Carboxyl-Kohlenstoff aus gezählt. Die Bezeichnung von Fettsäuren nach der Zahl der C-Atome und der Anzahl der Doppelbindungen hat sich als Kurzschreibweise eingebürgert (z. B. 16:0 für Palmitinsäure). Bei natürlich vorkommenden Fettsäuren können unterschiedliche Sättigungsgrade auftreten, wobei die Konfiguration, die Existenz oder Art einer trennenden Gruppe und Konjugationen z. B. mit einer Carbonylgruppe eine Rolle spielen können. Je mehr Doppelbindungen eine Fettsäure enthält, desto beweglicher ist sie. Der Sättigungsgrad der Fettsäuren einer Membran bestimmt deshalb deren Stabilität.
Andere Variationen der Fettsäurestruktur sind Ringe (Cyclopropan-, Cyclopentenring) oder Verzweigungen. Fettsäuren, die der Organismus nicht synthetisieren kann und mit der Nahrung aufnehmen muß, heißen essentielle Fettsäuren. Für den Menschen sind die Fettsäuren der Linolsäure-Gruppe essentiell.
Der Stoffwechsel der Fettsäuren
Aufnahme, Transport und Speicherung
Die Hydrolyse der Nahrungsfette erfolgt vor allem im Dünndarm. Sie werden zu Fettsäuren und Monoacylglycerin gespalten und von der Darmmucosa absorbiert, wo sie wieder zu Fetten aufgebaut werden. Dann werden sie als Chylomikronen – Fetttröpfchen mit netzartiger Proteinhülle – über das Lymphsystem in die Blutbahn abgegeben. Dort werden sie rasch hydrolysiert; die freigesetzten Fettsäuren werden vom Fettgewebe oder vom Albumin des Blutserums aufgenommen. Die übrigen Partikel werden in der Leber abgebaut. Die Leber kann aber auch Lipide synthetisieren. Diese werden an Proteine gebunden und als sogenannte Lipoproteine (z. B. Albumin) im Organismus verteilt (Karlson 1984). Zur Speicherung werden mehrere Fettsäuren an Glycerin gebunden und vor allem als Triacylglyceride (Triglyceride) gespeichert.
Biosynthese
Das Ausgangssubstrat für die Biosynthese von Fettsäuren ist Acetyl-CoA, das zumeist aus Citrat gebildet wird, wenn dieses aus den Mitochondrien in das Cytoplasma transportiert wird. Die Synthese findet im Cytoplasma statt. Der erste Syntheseschritt ist der Umbau des Acetyl-CoA zu Malonyl-CoA durch das Enzym Acetyl-CoA-Carboxylase (ACC). Das so aktivierte Molekül durchläuft mehrere Reaktionsschritte an einem Enzymkomplex. Die Reaktionen entsprechen einer Umkehrung der beta-Oxidation; pro Umlauf werden zwei C-Atome angefügt. Erreicht die Fettsäure eine Länge von 16 oder 18 C-Atomen wird sie vom Enzymkomplex abgelöst. Die Rate der Fettsäure-Synthese wird durch die Bereitstellung von Malonyl-CoA bestimmt (Thompson 1980).
Modifikationen
Die Zusammensetzung der Fettsäuren einer Art ist unter verschieden Bedingungen gleich; d. h. unabhängig von der Verfügbarkeit durch Nahrung oder der Biosynthese, die ja vor allem Palmitinsäure liefert. Somit müssen Organismen unterschiedliche Veränderungen an Fettsäuren vornehmen können. Im folgenden werden vor allem Mechanismen bei Eukaryoten beschrieben; Prokaryoten haben z. T. abweichende Mechanismen (Thompson 1980).
Verlängerung
Zur Verlängerung von Fettsäuren stehen bei Eukaryoten zwei Enzymsysteme zur Verfügung; in den Mitochondrien und in den Mikrosomen. In den Mitochondrien laufen zur Verlängerung einige der Reaktionen der β-Oxidation umgekehrt ab; den Abschluß bildet ein zusätzliches Enzym. In den Mitochondrien von Leber und Niere ist die Anwesenheit von NADH und NADPH nötig; in Herz- und Skelettmuskeln nur NADH. Allerdings wird das Enzymsystem von den potentiellen Hauptsubstraten (C16- und C18-Derivate) gehemmt, weshalb für die mitochondriale Fettsäureverlängerung andere Funktionen, wie die Regulation der Verfügbarkeit von Reduktionsequivalenten (NADH), vermutet wird.
Die mikrosomale Verlängerung stellt die Hauptquelle für längerkettige Fettsäuren (C18 bis C24) dar. Der Ablauf ist bisher unklar, aber vermutlich anders als in den Mitochondrien; es sind für jede Kettenlänge unterschiedliche Enzyme zur Verlängerung nötig.
Desaturierung
Die Fluidität der Membranen wird in erster Linie von dem Sättigungsgrad der enthaltenen Fettsäuren bestimmt. Das Einfügen einer Doppelbindung in die Kohlenstoff-Kette funktioniert in allen aeroben Organismen nach dem gleichen Prinzip. Es gibt jedoch Unterschiede in der Anzahl der Doppelbindungen und der Sequenz des Einfügens sowie spezifische Regulationsmechanismen. Die Verlängerung erfolgt an Acyl-Gruppen unterschiedlicher Verbindungen und ist die Übertragung eines Elektrons über eine Elektronentransportkette auf Sauerstoff.
Verzweigungen
Verzweigte Fettsäuren wie iso- und anteiso-Fettsäuren, die eine Methyl-Gruppe am Methylenkohlenstoff C2 bzw. C3 besitzen, entstehen, wenn die Fettsäuresynthese mit einem verzweigten Substrat statt Acetyl-CoA gestartet wird. Myobakterien können außerdem Methylgruppen an eine phospholipid-gebundene verzweigte Fettsäure anfügen.
Biohydrogenierung
Bei Bakterien, die im anaeroben Teil des Pansens von Wiederkäuern leben, können ungesättigte Fettsäuren des Pflanzenfetts mittels eines Flavin-Systems enzymatisch hydrogenieren.
Oxidation
Der oxidative Abbau von Fettsäuren vollzieht sich in den Mitochondrien der Eukaryoten und im Cytosol der Prokaryoten über eine gut bekannte Abfolge von Reaktionen. Der Abbau der aktivierten Fettsäuren in den Mitochondrien erfolgt über eine Folge von vier Reaktionen. Dabei werden jeweils C2-Bruchstücke als Acetyl-CoA abgespalten. Die Reaktionen betreffen das β-Atom, daher die Bezeichnung β-Oxidation. Das entstehende Acetyl-CoA wird im Citratzyklus vollständig zu CO2 oxidiert; der Wasserstoff wird auf FAD·H2 oder NAD+ übertragen und in der Atmungskette zu Wasser verbrannt, worauf FADH2 bzw. NAD+ oxidiert werden und wieder zur Verfügung stehen. Ungesättigte Fettsäuren durchlaufen die selben Schritte; ihre vollständige Oxidation erfordert jedoch die Aktion zweier zusätzlicher Enzyme.
Ein alternativer Mechanismus des Fettsäure-Katabolismus ist die α-Oxidation, die in den Mikrosomen stattfindet. Dabei werden freie Fettsäuren in Derivate freier Radikaler verwandelt; es entsteht eine Hyperoxy-Verbindung. Diese reagiert mit dem Derivat zu einer sehr instabilen d-α-Hyperoxy-Fettsäure, die entweder decarboxyliert und oxidiert werden kann, um eine um ein Atom verkürzte Fettsäure zu formen, oder sie wird zu einer d-α-Hydroxy-Fettsäure reduziert. Fettsäuren mit ungeraden Kettenlängen und α-Hydroxy-Fettsäuren findet man in Lipiden des Hirns.
Ebenfalls in den Mikrosomen findet die ω-Oxidation von Fettsäuren zu ω-Hydroxy-Fettsäuren statt. Diese werden schrittweise zu Dicarboxylsäuren konvertiert. Die Reaktion erfordert molekularen Sauerstoff und läuft unter Beteiligung des mikrosomalen Elektronentransportsystems ab. Es werden hauptsächlich Fettsäuren mittlerer Länge (C9 - C12) verarbeitet.