Vorlesung: Morphologie der Tiere, WS 2001/2002
Morphologie der Tiere - 3. Doppelstunde
Metazoa, Porifera, Cnidaria
Einführung in die Metazoa, die vielzelligen Tiere
Den einzelligen Tieren, den Protozoa, werden die vielzelligen Tiere, die Metazoa, gegenübergestellt. Alle Metazoa sind multizellulär, beweglich, heterotroph und durchlaufen während ihrer Embryonalentwicklung (Ontogenese) ein sogenanntes
Blastula-Stadium (von gr. blastos = Keim; deutsch als Blasenkeim bezeichnet).Bei den Metazoa unterscheidet man 29 Stämme. Einer dieser Stämme, Chordata, enthält die Vertebrata oder Wirbeltiere, zu denen auch der Mensch gehört. Alle anderen Stämme werden als Invertebraten bezeichnet (eine gebräuchliche Sammelbezeichnung für alle Tiere, die keine Wirbeltiere sind)!
Innerhalb der Metazoa wird noch weiter unterschieden zwischen den Parazoa (Porifera; s. u.), Placozoa (s. nächste Doppelstunde) und
Eumetazoa. Eumetazoa bedeutet "echte Vielzeller" und umfasst alle Tiere, die echte Epithelien haben (von gr. epi = auf, oder um; thele = wortwörtlich: die Brustwarze; im engeren Sinne: eine Zellschicht, die etwas umschliesst bzw. abdeckt).
Stamm Porifera (Schwämme oder Schwammtiere)
(von lat. porus = Pore, Öffnung und fere = tragen; also Porenträger)
Dieser Tierstamm enthält ca. 5000 rezente Arten (rezent = heute noch lebend). Davon leben ca. 150 im Süßwasser, alle anderen im Meer. Ihre Größe reicht von ca. 1 cm bis 2 m, es gibt viele Variationen in Form und Farbe (warum sie überhaupt farbig sind, ist unklar). Schwämme sind sicher mehrzellige Tiere (Metazoa), aber ihre Körperzellen sind noch nicht in Gewebe wie z. B. Epithelien organisiert. Deshalb werden sie als
Parazoa gegenüber den Eumetazoa abgegrenzt.Systematik (in Stichworten):
Das Phylum (Stamm) Porifera besteht aus 3 Klassen:
Calcarea (synonym: Calcispongiae; Kalkschwämme) Skelettnadeln aus Kalk (Calcit), kleine Formen, Flachwasser, kein Spongin
Hexactinellida (synonym: = Hyalospongiae; Glasschwämme oder auch Kieselschwämme); ca. 300 Gattungen, Skelettnadeln (Spiculae) aus Silikaten (Siliciumoxid). Triaxonale (= Skelettnadeln) haben 6 Spitzen (3 Achsen wie in einem Koordinatensystem) oder 4 (tetraxonale Spiculae, Spitzen weisen in die Ecken eines Tetraeders), 10-30 cm, einzeln stehend und von allen Formen die regelmäßigsten; besiedeln größere Meerestiefen (dominant z. B. in antarktischen Gewässern).
Demospongiae: (von gr. demos = Volk; Gemeinde, Kolonie (vgl. Begriffe wie Demokratie, Demoskopie); Hornschwämme) 90 % aller Arten, hier auch die 3 Familien der Süßwasserschwämme (z. B. der häufige Spongilla lacustris), Silikat-Skelette oder Spongin oder beides. Der Badeschwamm, Spongia officinalis, hat freundlicherweise keine Spiculae.
Bauplan:
Man unterscheidet bei Schwämmen 3 Bauplantypen:
Ascon-Typ
von gr. ascos = SchlauchSycon-Typ
von gr. sykon = FeigeLeucon-Typ
von gr. leucos = weiß; Leucon ist ein alterGattungsname (mit wahrscheinlich weißlichen
Schwämmen)
Diese Typen sind keine systematische Unterteilung!
Ascon- und Sycontyp gibt es nur bei Kalkschwämmen, den Leucon-Typ gibt es bei 95% aller Schwämme.
Schwämme besitzen bereits mehrere spezialisierte Zelltypen:
Pinakocyten (von gr. pinax = Teller, Brett, Tafel; hier wahrscheinlich eher im Sinne von Täfelung gemeint). Diese bilden das
Pinakoderm (eine Art Epidermis, aber ohne Basallamina, ohne Zellkontakte, also kein epithelialer Aufbau!)Choanocyten (Kragengeißelzellen; vgl. Choanoflagellata): Mit Microvillisaum am apikalen Ende um das Flagellum herum. Diese Zellen erzeugen durch ihren Cilienschlag Wasserströme und filtrieren die Nahrung durch ihre Microvilli-Reusen.
Sklerocyten: Stellen die Skelett-Nadeln her (Spiculae); sie liegen zusammen mit anderen, amoeboiden Zellen in der sogenannten Mesogloea (von gr. = mesos: mitten und ho gloios = klebriges Öl), auch Mesohyl genannt (von gr. hyalos = etwas Durchscheinendes, z. B. Glas). Gemeint ist damit die gallertige Mittelschicht, die leicht durchscheinend ist und die die Hauptmasse des Schwamms ausmacht.
Collencyten: stellen Kollagene her; bestimmte Collencyten, die Spongocyten, stellen Spongin her. Dieses ist ein kollagenähnliches Protein, das im Mesohyl stark vernetzt wird. Dieses bleibt beim Trocknen von Naturbadeschwämmen übrig.
Archaeocyten
Sowohl das Mesohyl mit dem Spongin oder anderen Komponenten, als auch die Spiculae, stellen spezielle Formen von extrazellulärer Matrix dar, wie letzte Stunde besprochen!
Lebensweise:
Schwämme sind Filtrierer. Der von den Choanocyten verursachte Wasserstrom gelangt durch sog.
Ostien (von lat. ostium = Mündung) in den Innenraum, der Spongocoel oder Atrium genannt wird, von dort hinaus durch eine größere Öffnung, das Osculum (von gr. os, oris = der Mund; Mündchen), wieder hinaus. Die Pumpleistung kann gewaltig sein: Ein Schwamm von 0.5 l Volumen, kann bis zu 2 l pro Minute umsetzen. Die Lebensweise bedingt, dass Schwämme klares Wasser bevorzugen (Verstopfung der Ostien!)Die Choanocyten filtrieren feinste Partikelchen aus dem Wasser, die durch Phagocytose verdaut werden. Oft kommen sog. Zooxanthellen (vgl. Protozoa) als Symbionten in der Mesogloea oder in spezialisierten amöboiden Zellen der Mesogloea vor. Als Symbionten kommen Dinoflagellaten (dann in non-motiler Form) oder Cyanobakterien vor. Bis 50-80% der vom Schwamm benötigten Energie kann von den photoautotrophen Symbionten kommen!
Fortpflanzung und Entwicklung:
Die Spermien entwickeln sich aus Choanocyten, die Eier aus Stammzellen der Mesogloea (Archaeocyten). Die Art der Befruchtung ist merkwürdig: Die Spermien werden von Choanocyten aufgenommen und zu den Eizellen transportiert. Als Larvenformen treten entweder eine Parenchymella oder eine Amphiblastula auf. Beide sind im Prinzip bewimperte Blastula-Stadien.
Bei Schwämmen gibt es erstmals die Andeutung einer Trennung zwischen Soma und Keimbahn: "sterbliche" somatische Zellen und "unsterbliche" Stammzellen (Roux-Weismann-Theorie), aber die Determination ist noch nicht vollständig. Choanocyten können sich noch in andere Zellentypen umwandeln.
Besonderheiten sind noch die Gemmulae: Dauerstadien zum Überwintern bei Süßwasserschwämmen. Im Prinzip sind diese ein Konglomerat aus Stammzellen, umhüllt mit Spiculae und einer Schutzhülle aus Spongin.
Wirtschaftliche Bedeutung:
Außer als Lieferanten von Badeschwämmen bekommen Schwämme zunehmend Bedeutung in der pharmakologischen Forschung.
Schwämme, insbesondere marine, sind bunt. Dies liegt nicht nur an eventuell vorhandenen Endosymbionten, sondern auch daran, dass amöboide Zellen der Mesogloea Pigmente synthetisieren und in Granula mit sich tragen. Die Bedeutung ist noch unklar, aber teilweise könnten es Warnfarben sein. Schwämme erzeugen nämlich eine ganze Reihe interessanter Substanzen, darunter Cytostatika (z. B. cyclische Peptide) und Toxine. Fischtoxische Substanzen dienen wahrscheinlich als Fraßschutz (bei manchen Meeresschildkröten, die offenbar immun gegen diese Gifte sind, bestehen 95% der Exkremente aus Spiculae!), Cytostatika und antibakteriell wirkende Substanzen dienen wahrscheinlich als Schutz gegen Bewuchs durch andere Organismen.
Näheres in: Müller, W. E. G; Schröder, H. C. (1997) Bioaktive Substanzen aus Schwämmen: Gene weisen den Weg bei der Suche nach neuen Arzneimitteln
Biologie in unserer Zeit, Band 27 (Nr. 6), Seiten 389-398
Evolution:
Die systematische Stellung der Porifera ist nicht hundertprozentig klar. Vor allem die Frage, ob die Schwämme die Stammform der Eumetazoa sind oder nicht, ist weiter offen.
- Die Larvalstadien gleichen dem Blastula-Stadium der Eumetazoa.
Einführung in die Eumetazoa
Wie oben bereits erwähnt, verfügen die Eumetazoa über
Epithelien. Epithelien sind spezielle Gewebe, die aus Zellen bestehen, die in einer Ebene fest miteinander verknüpft sind (die Zellen stehen in festem mechanischem, aber auch elektrischem Kontakt). Epithelien können dadurch verschiedene Kompartimente effektiv voneinander abgrenzen. Epithelzellen sind polarisiert (Polarisierung von Zellen ist aber bereits bei Protozoen sichtbar!), d. h. die Zellen haben ein sogenanntes apikales Ende (vom Körper oder Organ weg nach außen gerichtet) und ein basales Ende (dem Körper oder Organ zugewandt). Des weiteren sezernieren Epithelzellen auch Materialien als extrazelluläre Matrix (ECM). Am basalen Ende ist dies eine soganannte Basallamina (eine besonders dünne, aber mechanisch sehr stabile ECM). Am apikalen Ende kann die ECM z. B. eine Schleimschicht sein, aber es kann auch eine festere und komplexere ECM z. B. in Form einer Kalkschale oder Cuticula vorkommen.Wie diese Stichworte Schale und Cuticula andeuten, verfügen die Eumetazoa, insbesondere die größeren Formen, über
Stützgewebe. Besondere Formen des Stützgewebes sind Skelette. Man findet Innenskelette, Aussenskelette, Hydroskelette; harte und elastische Skelette.Ebenfalls mit der Größe korreliert die Art der Bewegung: Nur noch die Larven der Eumetazoa sind teilweise auf Bewegung durch Cilienschlag angewiesen.
Bewegungen werden ansonsten durch Muskeln erzeugt. Muskeln bestehen aus spezialisierten Zellen, in denen bestimmte Elemente des Cytoskeletts (Actin und bestimmte Myosine) zur Bewegungserzeugung besonders gut entwickelt sind. D. h. nicht, dass höhere Organismen die Cilien aufgegeben hätten (vgl. letzte Doppelstunde: Flimmerepithelien mit Cilien, Cilien als reizaufnehmende Strukturen in vielen Sinneszellen). Einige kleinere Vertreter der Eumetazoa (z. B. einige Plathelminthes (Plattwürmer) und Rotatorien (Rädertierchen)) können sich sowohl mit Cilien als auch mit Muskeln fortbewegen.Ähnlich wie Protozoen Nahrung in membranumschlossene Endocytosevesikel aufnehmen, besitzen Metazoen
von Epithelien umschlossene Gastralräume. Dadurch ist es möglich größere Nahrungsmengen oder größere Nahrungsteile (z. B. ganze andere Tiere oder Pflanzenteile) in einem abgeschlossenen Hohlraum "in Ruhe" mechanisch u. enzymatisch aufzuschließen, also im Prinzip wie in einer Nahrungsvakuole eines Einzellers, aber in viel größerem Maßstab. Wichtig ist, dass das Innere (Lumen) eines solchen Gastralraumes vom Körper aus betrachtet immer ein Teil der äußeren Umwelt bleibt! Auch der Inhalt unseres Magen-Darmtrakts ist ein Teil der Außenwelt!
Mit zunehmender Größe und Komplexität der Organismen entstehen Kommunikationsprobleme zwischen den Zellen. Deshalb haben sich im Lauf der Evolution wahrscheinlich
Nervensysteme und die Kommunikation mit Hormonen (humorales System) entwickeln müssen.Eine komplexe Struktur entsteht nicht von selbst. Für die
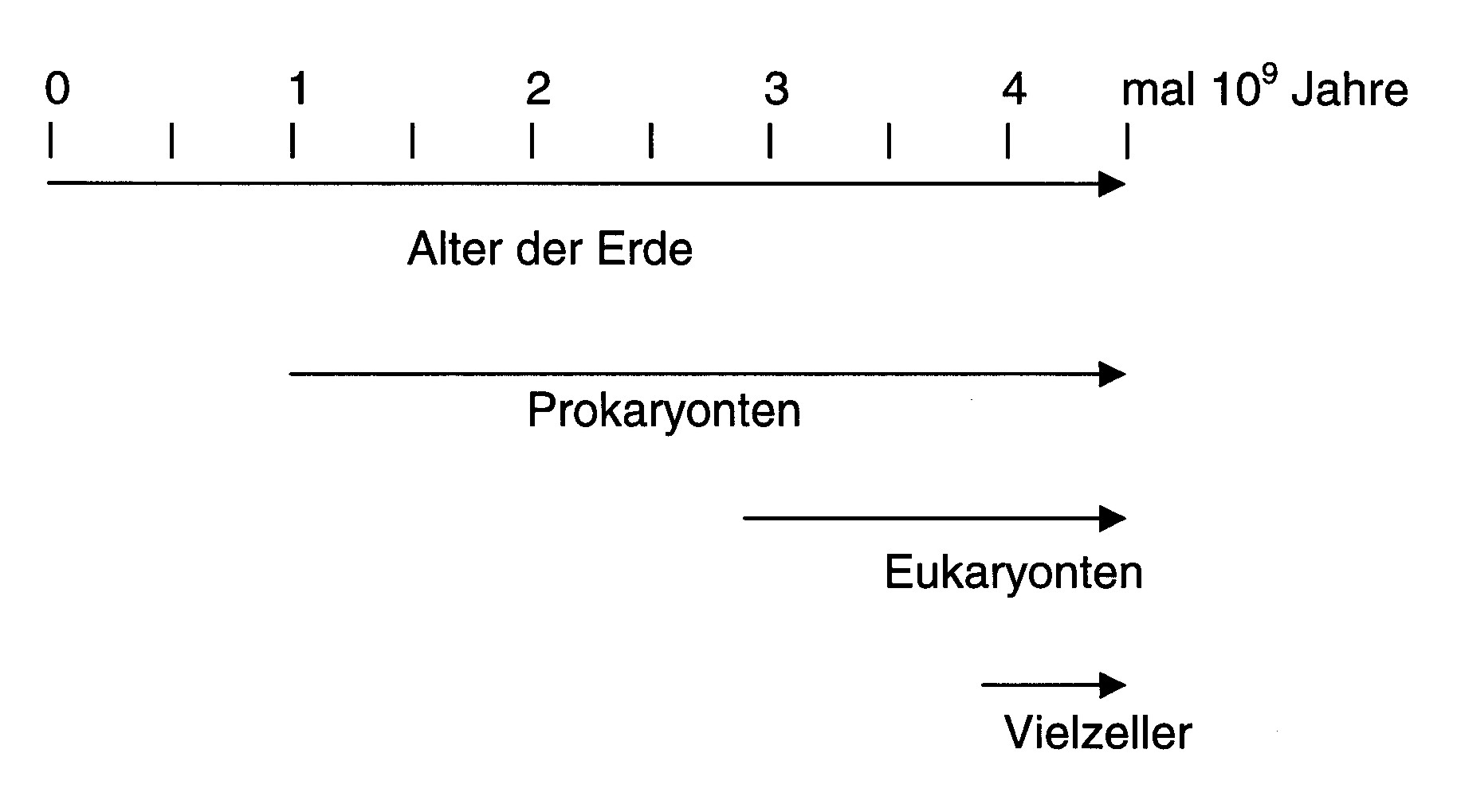
Embryonalentwicklung (Ontogenese) muss es ein Programm geben, dass aus undifferenzierten Stammzellen (Eizellen) spezialisierte Zellen, Gewebe, Organe und schließlich einen Organismus entstehen lässt. Auch hier ist die Kommunikation zwischen Zellen von entscheidender Bedeutung. Differenzierung heißt immer, dass Zellen bestimmte Gene anschalten und andere abschalten. Ein konzertiertes Programm muss ihnen sagen, wann und wo dies zu geschehen hat. Dies erfordert unter anderem ausgeklügelte Systeme des Informationsaustauschs und der Informationsverarbeitung. Zellen müssen ihr Verhalten miteinander koordinieren und die Einzelzelle muss im Gewebsverband wissen, welches ihre Position und Aufgabe ist. Multizelluläre Organismen sind ohne dies nicht denkbar und wahrscheinlich hat die Evolution von mehrzelligen Organismen deshalb so lange gedauert, weil diese komplexen Signalübertragungs- und -verarbeitungswege erst einmal evolvieren mussten (prokaryonte Bakterien gibt es schätzungsweise seit ca. 3.5 Milliarden Jahren, einzellige Eukaryonten seit ca. 1.5 und multizelluläre Organismen erst seit etwas mehr als 0.5 Milliarden Jahren!).
Stamm Cnidaria (von gr. knide = Nessel; Nesseltiere),
auch Coelenterata (von gr. hoilos = hohl u. enteron = Eingeweide; Hohltiere) genannt
Systematik (in Stichworten):
Der Stamm Cnidaria enthält alle Tiere, die landläufig als Polypen, Quallen, Seeanemonen und Korallen bekannt sind. Er umfasst
ca. 10000 Arten radiärsymmetrischer, vorwiegend marin lebender Tiere.
Es gibt zwei prinzipielle Erscheinungsformen:
Polyp (vegetativ) --> Medusen (geschlechtl.) --> Gameten (Eier u. Spermien) --> Polyp
Die Cnidaria werden in 3 Klassen unterteilt:
Klasse
Hydrozoa (von gr. hydor = Wasser u. zoon = Tier)2700 Arten, kleine Tiere (z. B. Bewuchs auf Hafenanlagen). Die wenigen Süßwasserarten gehören zu dieser Klasse. Entweder Polypen oder Medusen (oder Generationswechsel). Entweder getrenntgeschl. oder Zwitter. Kennzeichen: Keine Zellen in der Mesogloea, keine Nematocyten (Nesselzellen) im Entoderm, epidermale Gonaden.
Der bekannte einheimische Süßwasserpolyp Hydra lebt. z. B. solitär, zeigt aber vegetative Vermehrung. Die überwiegende Mehrheit der Hydrozoa bildet Kolonien. Koloniebildung bedingt oft Skelettbildung zur Verstärkung (meist Chitin, selten Kalk), manchmal tritt auch Arbeitsteilung zwischen verschiedenen Polypentypen auf. Staatsquallen (Siphonophora) sind dann das Beispiel wo alle Morphen sozusagen sinnvolle Teilaufgaben bekommen: Als Schwimmglocke dient eine Meduse, verschiedene Polypen dienen zum Fressen und zur Verteidigung, schließlich gibt es kleine Fortpflanzungsmedusen.
Auch andere Hydrozoen formen Medusen-Knospen. Diese können sich zu freischwimmenden Formen entwickeln, können aber auch an der Kolonie verbleiben. In jedem Fall entwickeln sich in den Medusen-Formen die Gonaden (Gonophoren). Wenn die Medusen nur von bestimmten Polypen geformt werden, dann nennt man diese Gonozoide. Die Medusen der Hydrozoa sind klein.
Klasse
Scyphozoa, Cubozoa (gr. scyphos = Becher, Trichter; lat. cubus = Würfel). Die Cubozoa werden in manchen Systematiken als getrennte Klasse geführt.Alles was gemeinhin als Qualle bezeichnet wird. Es gibt ca. 200 Arten Scyphozoen, 15 Arten Cubozoen. Größer als Hydromedusen (2 - 40 cm, maximal 2 m). Gefährlich aufgrund ihrer Toxine, einige australische Spezies können tödlich sein. In der Mesogloea befinden sich amöboide Zellen. Getrenntgeschlechtlich, Gonaden am Gastroderm (beidseitig der Septen (Trennwände), wenn diese vorhanden), aus den Larven entstehen kleine Polypen ähnlich Hydra, aber mit Septen im Gastralraum. Diese können sich in eine Meduse umwandeln, oder es enstehen kleine Medusen am oralen Ende (Strobilation; gr. strobilos = Wirbel, Zapfen). Medusenformen haben oft komliziertere Nervensystem und Sinnesorgane (Photorezeptoren, Ocellen, Statocysten) als Polypenformen.
Klasse Anthozoa (von gr. anthos = Blume)
6000 Arten, alles Polypen, keinerlei Medusenstadium, septierter Gastralraum, Gonaden im Gastroderm, Zellen in der Mesogloea, Nematocyten ohne Operculum (von lat. Deckel), Nematocyten mit Klebfäden hauptsächlich in den Tentakeln, ernähren sich z. T. von großer Beute (Fische, Krabben), hermaphroditisch
Werden weiter unterteilt in
Hexacorallia und Octocorallia. Anthozoen mit 12 Tentakeln und Septen (oder mehrfaches davon) heißen Hexacorallia. Dazu gehören z. B. die Skleraktinia (Steinkorallen) und Actiniaria (Seeanemonen). Octocorallia haben 8 Tentakel, die gefiedert sind, und immer 8 komplette Septen. Manche haben ein Innenskelett aus teilweise verkalkter Mesogloea. Hierzu gehören z. B. auch die Hornkorallen (Gorgonacea), und dazu wiederum die bekannten roten Korallen (Korallenschmuck).Übersicht:
|
Hydrozoa |
Scyphozoa |
Anthozoa |
|
|
Septen im Gastralraum der Polypen |
- |
+ |
+ |
|
Zellen in der Mesogloea |
- |
+ |
+ |
|
Gonaden |
ectodermal |
gastrodermal |
gastrodermal |
|
Generationswechsel |
Polyp - Meduse |
Polyp - Meduse |
nur Polypen |
|
Geschlechter |
m. + f. oder Zwitter |
m. + f. |
Zwitter |
Bauplan:
Cnidaria sind
radiärsymmetrisch gebaut. Diese Radiärsymmetrie wird in den Lehrbüchern oft als ursprünglich oder primitiv angesprochen. Für die sessile Lebensweise ist diese Symmetrie jedoch von Vorteil. Viele Vertreter anderer Tierstämme sind mit der sessilen Lebensweise wieder ganz oder ansatzweise zur Radiärsymmetrie übergegangen.Der Körper der Cnidaria besteht im Prinzip aus einem zumeist zylindrischen Hohlraum (Gastralraum, Coelenteron), der von zwei echten Epithelschichten umschlossen wird: Ein inneres Epithel, das
Entoderm oder Gastroderm, und ein äußeres Epithel, das Ectoderm (von gr. entos = innen, bzw. ektos = außen, derma = Haut und gaster = Magen, Bauch). Zwischen diesen befindet sich die Mesogloea in mehr oder weniger starker Ausprägung.Bei vielen Hydrozoen ist die Mesogloea nicht mehr als die vereinigte Basallamina der beiden Epithelien. Bei anderen Cnidaria, besonders bei den Scyphozoa, ist sie eine komplexe und dicke Bindegewebsschicht (mit oder ohne Bindegewebszellen) und gibt ein Beispiel für ein elastisches Skelett. Das englische Wort für Qualle, jellyfish, beschreibt diesen Zustand treffend!
Am oralen Ende (das Ende, wo die Mundöffnung liegt) bilden sich Tentakel (von lat. tentaculum = Fühler) aus. Das Coelenteron kann durch Septen unterteilt sein. Bei Medusen kann die Einengung des ursprünglichen Hohlraums durch Septen so weit gehen, dass teilweise nur Kanäle übrig bleiben (meist Ringkanäle). Dies macht deutlich, dass der Gastralraum nicht nur Magen-Darm-Funktion hat, sondern auch zur Verteilung der Nährstoffe wichtig ist. Letztere Funktion wird bei anderen Metazoen zumeist durch das Blutgefäßsystem ausgeübt.
Zelltypen in den Epithelien:
Der Epithel-Aufbau kann als primitiv im Sinne von "ursprünglich" angesehen werden. Andererseits haben die Cnidaria aber einen Zelltyp hervorgebracht, der an Komplexität durch fast nichts zu überbieten ist, die Nesselzellen (s. u.).
Ectoderm und Entoderm setzen sich in der Hauptsache aus
Myoepithelzellen (synonym: Epithelmuskelzellen) zusammen (von gr. mys, myos = Maus, Muskel). Diese sind eine Art Mischtyp aus Epithelzellen und Muskelzellen. Im Ectoderm sind die kontraktilen Fasern in Körperlängsrichtung orientiert, im Entoderm liegen sie quer zur Längsachse des Tieres und fungieren so als Ringmuskulatur. Im Entoderm tragen sie außerdem Cilien und können Zooxanthellen (bei Korallen hauptsächlich) enthalten.Reine Muskelzellen sind im Lauf der Evolution wahrscheinlich dadurch entstanden, dass bei solchen Zellen die Epithelfunktion verloren ging. Epithelmuskelzellen gibt es nicht nur bei Cnidariern sondern z. B. auch in den Schweiß- und Milchdrüsen und in der Iris von Wirbeltieren. Die Epithelmuskelzellen bilden bereits spezialisierte Zellkontaktstrukturen wie adhesion belts und gap junctions (letztere nur bei Hydrozoen) aus, die man auch bei den Epithelzellen höherer Metazoen findet (näheres siehe Lehrbücher zur Zellbiologie).
Neben den Myoepithelzellen treten auf:
Die (namensgebenden)
Cnidocyten oder Nematocyten (von gr. nema = Faden) liegen oft nur im Ectoderm. Sie sind wichtig für Beutefang und Verteidigung (und für Zoologen f. d. Bestimmung der Arten), sonst siehe unten im Detail.Sekretorische Zellen
(Drüsenzellen): Wichtige Aufgaben sind die Produktion von Schleim, der die Auslösung der Nesselzellen verhindert, wenn sich das Tier selbst berührt. Weiter spielen sie eine Rolle beim Anheften ans Substrat, beim Bau von Skelettelementen (Korallen) und, im Gastroderm, bei der Sekretion von Verdauungsenzymen.Sinneszellen
(synonym: Rezeptorzellen) und Nervenzellen: Die Sinneszellen haben Cilien, die Nervenzellen liegen nahe an der Mesogloea und bilden diffuse Nervennetze. Es gibt offenbar keine "klassischen" Transmitter (wie z. B. Acetylcholin), als Transmitter dienen Peptide.Interstitialzellen
: liegen in der Mesogloea und stellen Stammzellen für alle Zelltypen dar, inklusive der Keimzellen. Manche Cnidaria sind Regenerationskünstler!
Exkurs: Die Nesselzellen
Die
Nematocyten (synonym: Cniden oder Cnidocyten; Nesselzellen) sind mit die komplexesten Zellen, die es im Tierreich gibt. Nesselzellen sind sensorische Zellen (mit sensorischem Cilium, hier Cnidocil genannt) mit einem großen spezialisierten Sekretionsvesikel (die Nesselkapsel; 5-100 µm groß). Es gibt ungefähr 20-30 verschieden Formen von Nesselzellen, die dann je nach ihrer Funktion verschiedene Namen haben (Volventen, Glutinanten, Penetranten etc.).Die Nesselkapsel ist im Prinzip ein riesiger
Exocytosevesikel. Dieser ist von einem Korb aus Microtubuli und Intermediärfilamenten umgeben. Diese Fasern stehen mit Actinfasern um die Stereocilien (die heißen dummerweise so, aber es sind keine Cilien, sondern Microvilli!) um das Cnidocil in Verbindung. Die Entladung erfolgt innerhalb von 4 ms; die eigentliche Ausstülpung der Kapsel innerhalb von 10 µs und mit 40000 g (g=Erdbeschleunigung)!Die äußere Kapselwand besteht aus vielen Proteinen, die innere Kapselwand aus einem Minikollagen. Diese Proteine bilden eine Netzstruktur, die dem enormen Innendruck standhalten muss (bis 150 Bar, d. h. das 100fache eines Autoreifens!) Der Innendruck wird osmotisch erzeugt durch Anhäufung von Polyanionen (Poly-gamma-glutamate, gibt es sonst nur bei Bakterien). Die Centriolen wirken bei der Formgebung der Nesselkapsel mit. Der Schlauch ist zunächst ausserhalb der Kapsel und die Mechanismen der Einstülpung, der Stilett-Bildung und des Tranfers von Polyanionen in die Kapsel ist noch weitgehend unklar.
Kapselflüssigkeit enhält noch ca. 30 unidentifizierte Proteine, davon sind einige Toxine, aber es gibt (zumindest bei Hydra,wo dies untersucht wurde) keine Chitinasen, Kollagenasen, und Proteasen, also keine Verdauungshilfen. Die Toxine der Seeanemonen sind am besten untersucht; es handelt sich um sehr potente Nervengifte.
Generation: von den ca. 100000 Zellen einer Hydra sind ca. 25000 reife Nematocyten und 40000 sind heranreifende Nematocyten (Der kleine Süßwasserpolyp ist also eine Nesselzell-Fabrik!). Nematocyten werden von Stammzell-Nestern im Rumpf gebildet und wandern erst später in die Tentakel.
Näheres bei: Müller, W. A. (1996) Pattern formation in the immortal Hydra. Trends. Genet. 12:91-96 - Tardent, P. (1995) The cnidarian cnidocyte, a high-tech cellular weaponry. BioEssays 17:351-362 - Holstein, T. W. (1995) Nematocyten. Biologie in unserer Zeit 25:161-169
Fortpflanzung und Entwicklung:
Wie schon erwähnt sind die Medusenformen im allgemeinen die Geschlechtsformen. Aus befruchteten Eiern entwickeln sich sogenannte
Planulalarven. Oft gibt es vegetative Vermehrung durch Knospen. Aus dieser Art Vermehrung ist wahrscheinlich auch die bei Cnidariern stark ausgeprägte Tendenz zur Bildung von Kolonien zu erklären.
Wirtschaftliche Bedeutung:
Die direkt Bedeutung ist klein (Schmuckindustrie, teilweise pharmakologische Forschung (Toxine)), die indirekte um so größer. Korallenriffe sind oft natürlicher Küstenschutz und zudem Tourismusziele. Die Riffe haben auch immense Bedeutung für die Meeresökologie.